
Diet-Tissue Dynamics of EFA Nutrients
Omega-3 and omega-6 essential fatty acids (EFA) were first recognized as needed vitamin-like nutrients in 1929. The similar chemistry and metabolism of n-3 linolenic and n-6 linoleic acid was clarified during 1930 to 1960. The n-3 and n-6 types compete for elongation, desaturation and esterification steps as they efficiently form HUFA that accumulate in tissue phospholipids. As a result, eating more of one type lowers the proportion of the other in tissue HUFA.
Early signs of the need for EFA were poor growth and skin dysfunction. Probably less than 0.5% food energy as n-6 linoleic acid prevents deficiency symptoms in human infants. In 1964, researchers saw that n-3 and n-6 HUFA form various prostaglandins. That opened a new area of dramatically different actions of EFA in human physiology. We now know that dietary intakes of n-6 linoleic acid causing the %n-6 in HUFA to be above 50% are associated with higher risks for health disorders. Too much omega-6 creates a transition (4:53 min video) between healthy physiology and pathophysiology with excessive immune-inflammatory events.
Health risk assessment (1 min.video) can monitor HUFA balance (the %n-6 in HUFA) that results from food choices. What people eat is important. With no dietary omega-3 nutrients, the dietary omega-6 linoleate has a very narrow therapeutic window that is made wider by eating omega-3 nutrients. Eating fewer omega-6 nutrients makes the competing omega-3 nutrients more effective in diminishing risk for chronic inflammatory processes. Benefits in balancing omega-3 and omega-6 HUFA are discussed in Video-1 (4.5 min.video) and Video-2 (7 min.video).
Pioneering 1963 work of Mohrhauer and Holman (1 min.video) showed potent competition of n-3 and n-6 nutrients in forming tissue HUFA . Some newer knowledge is noted below.
ACYL CHAIN ACCUMULATION AND HIGH-THROUGHPUT HUFA ASSAYS
The proportions of omega-3 (n-3) and omega-6 (n-6) in 20- and 22-carbon highly unsaturated fatty acids with 3 or more double bonds (HUFA) accumulated in tissue (e.g., the %n-6 in HUFA) reflect intakes of n-6 and n-3 fatty acids. An empirical equation (the Lands’ Equation) was developed previously to use dietary intakes of n-6 and n-3 HUFA and their 18-carbon precursors to predict the %n-6 in HUFA of humans. Linear regression analyses comparing observed and predicted %n-6 in HUFA had a correlation of 0.73 (p<0.000000) for nearly 4,000 individuals in 34 studies with 92 subject groups from 11 different countries. Strandjord SE, Lands B, Hibbeln JR. Validation of an equation predicting highly unsaturated fatty acid (HUFA) compositions of human blood fractions from dietary intakes of both HUFAs and their precursors. Prostaglandins Leukot Essent Fatty Acids. 2017 Mar 8. pii: S0952-3278(16)30157-0. doi: 10.1016/j.plefa.2017.03.005. PMID: 28390839
Relative dietary supplies of competing n-3 and n-6 nutrients determine the balance of 20- and 22-carbon n-3 and n-6 highly unsaturated fatty acids (HUFA) that accumulate competitively at the 2-position of tissue phospholipids. In turn, the HUFA balance (%n-6 in HUFA) affects the likely intensity of n-6 eicosanoid actions. As a result, measures of HUFA balance are important, valid biomarkers for designing and monitoring successful preventive nutrition interventions. Successful interventions must also consider how fatty acid ligands saturate binding sites of enzymes and receptors and give paradoxical dose-response results. Lands B, Bibus D, Stark KD. Dynamic interactions of n-3 and n-6 fatty acid nutrients. Prostaglandins Leukot Essent Fatty Acids. 2017 Jan 25. pii: S0952-3278(16)30152-1. doi: 10.1016/j.plefa.2017.01.012. PMID: 28189338
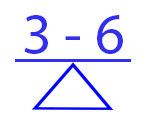
The impact of a food item upon the useful health risk assessment biomarker, HUFA balance, is easily evident in the Omega 3-6 Balance Food Score that summarizes in a single value the balance among eleven omega-3 and omega-6 essential fatty acids in a food. Foods with more positive Omega 3-6 Balance Food Scores will increase the percent of omega-3 in the biomarker, whereas those with more negative Scores will increase the percent of omega-6 in the biomarker. Lands B, Lamoreaux E. Using 3-6 differences in essential fatty acids rather than 3/6 ratios gives useful food balance scores. Nutr Metab (Lond). 2012 May 24;9(1):46. doi: 10.1186/1743-7075-9-46. PMID: 22624598
Two basic concepts learned during my graduate studies are embedded in all that I did and do: “Connect the Dots” and “Make Quantitative Models of Reality”. Lands B. Everything is connected to everything else. J Biol Chem. 2011 Dec 23;286(51):43589-95. doi: 10.1074/jbc.X111.318873 PMID: 22045809
All good research begins with wishful thinking about ‘Why things happen’ before scientific discipline converts it into controlled interventions that address `How does this happen?’ and `What is the consequence when this happens?’. This review examines some selective acyl chain actions. Lands WE. Stories about acyl chains. Biochim Biophys Acta. 2000 Jan 3;1483(1):1-14. PMID: 10601692
Similar selectivities for metabolism in mice, rats, and humans permit data from all three to be useful in estimating long-term tissue responses to different diets. Tissue phospholipids have average proportions of n-3 and n-6 in HUFA related hyperbolically to dietary supplies of the precursors, linoleate (18:2n-6) and linolenate (18:3n-3). In contrast, tissue triacylglycerols accumulate the precursors in a linear relation to their dietary supply. Lands WE. Long-term fat intake and biomarkers. Am J Clin Nutr. 1995 Mar;61(3 Suppl):721S-725S. PMID: 7879743
Competition between n-3 and n-6 HUFA can diminish the proportion of n-6 in eicosanoid precursors of a tissue. That diminishes the intensity of tissue responses mediated by n-6 eicosanoids. The proportions of n-3 and n-6 HUFA in human plasma relate to the dietary intake of 18:3 n-3 and 18:2 n-6 by an empirical hyperbolic equation very similar to the equation reported for laboratory rats. The HUFA balance may be useful in estimating a diet-related tendency for severe thrombotic, inflammatory or other disorders mediated by n-6 eicosanoids. Additional constants and terms were included in the equations to account for effects of dietary n-3 HUFA. A smaller constant for n-3 HUFA confirmed a greater competitive effectiveness of n-3 HUFA compared to 18:3n-3 in decreasing the 20:4n-6 in tissue lipids. The equations successfully used fatty acid contents of plasma lipids to estimate daily intakes of supplemental n-3 HUFA. Lands WE, Libelt B, Morris A, Kramer NC, Prewitt TE, Bowen P, Schmeisser D, Davidson MH, Burns JH. Maintenance of lower proportions of n-6 eicosanoid precursors in phospholipids of human plasma in response to added dietary n-3 fatty acids. Biochim Biophys Acta. 1992 Dec 10;1180(2):147-62. PMID: 1463766
A method for combining data on fatty acid composition into subsets describes quantitative competitive selectivities in metabolic and transport events that maintain fatty acid compositions in tissue lipids. Tissue triglycerides maintained a linear relationship with 18-carbon polyunsaturated fatty acids in the diet. Phospholipids in liver, plasma and red blood cells maintained about 45% of the fatty acids as saturated fatty acids and 20-30% as HUFA, irrespective of very different proportions of n-3, n-6 and n-9 types of fatty acids. The n-3, n-6 and n-9 HUFA were maintained in a competitive hyperbolic relationship with apparent EC50 values near 0.1% of dietary calories for 18:2n-6 and 18:3n-3. A focus on HUFA balance may be useful in predicting the influence of diet upon tissue levels of substrates and antagonists of eicosanoid biosynthesis. Lands WE, Morris A, Libelt B. Quantitative effects of dietary polyunsaturated fats on the composition of fatty acids in rat tissues. Lipids. 1990 Sep;25(9):505-16. PMID: 2250586
Polyunsaturated fatty acids are transferred from their coenzyme A thiol esters to phospholipid acceptors with very different maximal velocities. The low apparent Km values for the thiol esters in acylating 1-acylglycerol-3-phosphocholine were estimated relative to that for arachidonyl-CoA. Acyl-CoA esters of saturated acids, as well as those for 22:1, 22:2, and 22:3, had negligible ability to compete for the active sites of the 1-acyl-GPC acyltransferase system. The acyl-CoA esters of arachidonate (20:4n-6), eicosatrienoate (20:3n-6), eicosapentaenoate (20:5n-3), and both isomers of linolenate (18:3n-6 and n-3) were preferentially used by the 1-acyl-GPC acyltransferases. The system from liver has a high selectivity for unsaturated acids but does not appear to discriminate between n-6 and n-3 acids that are precursors of prostaglandins and leukotrienes. Lands WE, Inoue M, Sugiura Y, Okuyama H. Selective incorporation of polyunsaturated fatty acids into phosphatidylcholine by rat liver microsomes. J Biol Chem. 1982 Dec 25;257(24):14968-72. PMID: 7174678
The in vitro selectivity of microsomal acyl-CoA:monoacyl- sn-glycerol-3-phosphate acyltransferases varied depending upon the incubation conditions. Higher selectivity for linoleate occurs at lower monoacyl-sn-glycerol-3-phosphate concentrations. At very low I-acyl-GP concentrations, linoleate tended to exclude palmitate and arachidonate from the 2-position of diacyl-GP, whereas high 1-acyl-GP concentrations gave relatively nonselective acylation. Thus, a low in vivo steady-state level of acyl-GP may explain preferential accumulation of linoleate (18:2n-6) in triglycerides. Okuyama H, Lands WE. Variable selectivities of acyl coenzyme A:monoacylglycerophosphate acyltransferases in rat liver. J Biol Chem. 1972 Mar 10;247(5):1414-23. PMID: 5012315
The synthesis of diglyceride and triglyceride species from 14C-glycerol was examined in rat liver slices. 70% of newly synthesized diglycerides contained one or two double bonds per molecule, suggesting a preference for linoleate and oleate in the de novo pathway. Hill EE, Lands WE, Slakey SP. The incorporation of 14C-glycerol into different species of diglycerides and triglycerides in rat liver slices. Lipids. 1968 Sep;3(5):411-6. PMID: 17805819
Human erythrocyte acyltransferases are likely significant in controlling the fatty acid composition at the 2-position of lecithin. Waku K, Lands WE. Control of lecithin biosynthesis in erythrocyte membranes. J Lipid Res. 1968 Jan;9(1):12-8. PMID: 5637422
Acyltransferase rates for 66 combinations of substrates (6 acylglycerolphosphorylcholines and II acylcoenzyme A esters) were measured using a preparation from pig liver. The position being acylated had more influence than the composition of fatty acids in the 1-acylglycerol phosphorylcholine. Brandt AE, Lands WE. The effect of acyl-group composition on the rate of acyltransferase-catalyzed synthesis of lecithin. Biochim Biophys Acta. 1967 Dec 5;144(3):605-12. PMID: 5624813
The acyl-CoA:acyl-GPC acyltransferases have a marked specificity for both the position to be acylated and the particular acylCoA involved. In some cases, observed distributions of fatty acids in naturally occurring lecithins and triglycerides are similar to distributions predicted from in vitro studies of acyltransferase activity. Lands WE, Hart P. The control of fatty acid composition in glycerolipids. J Am Oil Chem Soc. 1966 May;43(5):290-5. PMID: 5933464
HIGH-THROUGHPUT HUFA ASSAYS
Large population studies have been hampered by inadequate high-throughput analytical technologies. An automated high throughput fatty acid analysis was developed using direct transesterification and automation of chemical procedures, data acquisition and automatic data processing. This is the first report of a complete, validated, cost-effective, automated, high-throughput fatty acid measurement method along with application to a population-based study. Lin YH, Salem N Jr, Wells EM, Zhou W, Loewke JD, Brown JA, Lands WE, Goldman LR, Hibbeln JR. Automated high-throughput fatty acid analysis of umbilical cord serum and application to an epidemiological study. Lipids. 2012 May;47(5):527-39. PMID: 22430941
A simple blood assay uses whole blood, collected by finger prick, stored on absorbant paper, subjected to direct methylation and quantified by automated GC. The assay provides a non-invasive, rapid and reliable method of HUFA quantification with the %n-3 in HUFA value providing a useful biomarker for large-scale nutritional trials. Bell JG, Mackinlay EE, Dick JR, Younger I, Lands B, Gilhooly T. Using a fingertip whole blood sample for rapid fatty acid measurement: method validation and correlation with erythrocyte polar lipid compositions in UK subjects. Br J Nutr. 2011 Nov;106(9):1408-15. PMID: 21736805
A simple method for analyzing fatty acids in a 50 microliter finger-tip blood samples from an individual may be useful in wide-scale screening of different individuals. Ohta A, Mayo MC, Kramer N, Lands WE. Rapid analysis of fatty acids in plasma lipids. Lipids. 1990 Nov;25(11):742-7. PMID: 2280679
Updated November, 2017